Extrait du B.O.
Dans les cellules, aussi bien autotrophes qu’hétérotrophes, la matière organique a trois devenirs : elle peut (1) être stockée ou exportée, (2) servir à la biosynthèse de nouvelles molécules organiques, ou (3) entrer dans les voies cataboliques fournissant son énergie à la cellule.
(1) Le stockage de la matière organique permet de constituer des réserves. Chez les Chlorophytes, les trioses phosphates produits par le cycle de Calvin sont stockés transitoirement sous forme d’amidon dans le stroma chloroplastique ou sont convertis en glucides exportables vers d’autres cellules. Le glucose absorbé par une cellule hétérotrophe animale peut être stocké sous forme de glycogène dans le cytosol. Le stockage ou le déstockage des réserves glucidiques dépend de l’activité d’enzymes.
(2) La matière organique permet de synthétiser de nouvelles molécules : c’est l’anabolisme. Les principales molécules du vivant ont un squelette carboné qui dérive d’intermédiaires de voies du métabolisme. Des interconversions entre familles de molécules sont possibles, elles aboutissent à la biosynthèse des principales molécules à rôle structural, métabolique ou informationnel à partir de carrefours métaboliques.
(3) La matière organique absorbée ou stockée peut entrer dans les voies cataboliques desquelles la cellule tire son énergie. Le catabolisme peut être oxydatif aérobie (respiration) ou non (fermentation).
La glycolyse est une voie métabolique permettant la biosynthèse d’ATP (par transphosphorylation ou phosphorylation liée au substrat), de coenzymes réduits et de pyruvate par une chaîne de réactions partant du glucose. L’oxydation du glycéraldéhyde-3-P dans le cytosol en est une réaction clef. La glycolyse est l’objet d’un contrôle cellulaire. Il participe à l’ajustement de la production d’ATP aux besoins de la cellule.
Dans le cas de la fermentation, la glycolyse est la seule voie de production d’ATP. Des réactions biochimiques spécifiques de chaque voie fermentaire oxydent les coenzymes réduits, les rendant à nouveau disponibles pour la glycolyse.
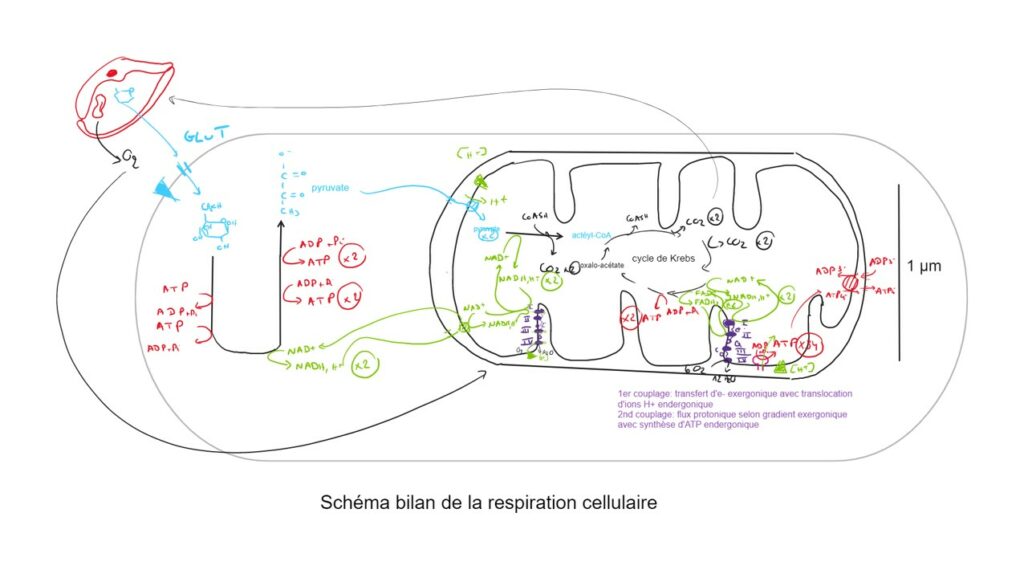
Dans le cas de la respiration cellulaire, le pyruvate est importé dans la mitochondrie et subit une décarboxylation à l’origine d’acétyl-CoA.
Les acides gras peuvent également être importés dans la matrice mitochondriale et produire de l’acétyl-CoA par β-oxydation. Le catabolisme des acides aminés passe par le pyruvate ou un intermédiaire du cycle de Krebs.
Le cycle de Krebs est une voie de convergence du catabolisme utilisant l’acétyl-CoA chez toutes les cellules à catabolisme aérobie. Il réalise la décarboxylation oxydative totale des composés, couplée à la production de nucléotides énergétiques et la réduction de coenzymes.
La chaîne respiratoire est une chaîne de transfert d’électrons issus de coenzymes réduits vers un accepteur final minéral à plus fort potentiel d’oxydoréduction. Ce transfert est associé à des conversions chimio-osmotiques (via la chaîne respiratoire) et osmo-chimiques (via l’ATP synthase) qui permettent la production d’ATP. L’ATP est donc synthétisée en quantité variable selon le métabolite initial et la voie métabolique.
Documents
-
 SV-E-2-devenir_matiere_organique_planches_2026
SV-E-2-devenir_matiere_organique_planches_2026
Taille: 9 Mo Téléchargements: 150